Экологическая модель растительного покрова степей бассейна Дона
Аннотация
Дата поступления статьи: 30.11.2013В работе приведены результаты ординационного анализа растительного покрова степей бассейна Дона
Ключевые слова: ординация, бассейн Дона, степи
05.13.18 - Математическое моделирование, численные методы и комплексы программ
Введение
Наука, как коллективное творчество ученых и как результат их взаимодействия с природой, определяет информационное развитие социума. В связи с этим, в современной схеме научного познания мира особо важным разделом является разработка моделей его организации [1]. С середины XX века ботаническая наука взяла направление от описательного подхода в сторону объяснения существующих закономерностей в растительном покрове [2] и неотъемлемой частью их познания стало построение математических моделей. Изучение экологии растительных сообществ постепенно стало осуществляться с учетом особенностей и факторов, определяющих поведение растительных объектов, причем, по характеру получаемых результатов, они уже приближались к экспериментальному исследованию, а не к простой статистической обработке данных [3].
С появлением компьютерной техники, разнообразных программных комплексов и возможности автоматизации вычислений, ординация стала необходимым инструментом в познании экологических закономерностей распределения растительных сообществ [4]. В экологии сообществ, в исследованиях структуры растительного покрова стал применяться анализ изменения различных показателей в пространстве экологических факторов, которые интерпретировались как комплексные градиенты среды. Появилась возможность построения таких моделей растительного покрова, при которых изучалась вариабельность растительных объектов на осях экологических факторов, или на осях ординации, выполнялся анализ положения растительных сообществ в экологическом пространстве.
Ординация рассматривается как совокупность многомерных методов обработки большого объема данных о связи растительности и условий среды. На данный период существует две группы методов ординации: прямая и непрямая. Прямая ординация показывает изменение видового состава вдоль выбранных конкретных экологических факторов. В отдельных случаях применяются «полупрямые» методы (ось фактора среды строится на основе знания его максимума и минимума).
Непрямая ординация позволяет определить экологически значимые оси максимального варьирования (с наибольшей нагрузкой), отражающие комплексные градиенты лимитирующих факторов среды; выявить важнейшие экологические факторы без их прямой оценки, интерпретируя оси максимальных нагрузок, или оси ординации. Поэтому выявление связи между растительностью и средой с применением методов непрямой ординации, в которых используются репрезентативные данные о видовом составе геоботанических описаний, теперь признается важнейшей задачей любого фитоценологического исследования [5].
Растительный покров выражен на определенной территории, поэтому пространственное моделирование как особая географическая форма моделирования, которая дополняет функциональную модель растительного сообщества, было особенно важно при построении экологических моделей.
Основной целью наших исследований являлось выявление экологических закономерностей распределения растительного покрова степей бассейна Дона на основе построения экологических моделей с использованием методов непрямой ординации и картографической модели растительности как географической формы моделирования.
Материалы и методы
В течение длительного времени в бассейне Дона, в пределах исследуемой территории Ростовской области, наблюдалось стихийное использование природных ресурсов и поэтому крупные массивы степей в настоящее время оказались практически уничтоженными. Господствуя в прошлом и занимая около 90 % территории Ростовской области, степи подверглись тотальной распашке, а сохранившиеся фрагментами степные участки составляют теперь всего 16,6 – 17,3 % от ее общей площади [6, 7]. Эти степи используются в качестве природных кормовых угодий, находятся на различных стадиях пасквальной дигрессии и часто преобразованы деятельностью человека, представляя антропогенные модификации степной растительности – «полуприродные» сообщества с большой долей сорных и чужеродных видов. Несмотря на это, здесь, на границе Причерноморских и Заволжско-Казахстанских степей [8, 9], в ключевом для сохранения биоразнообразия степей регионе, сохранились уникальные сообщества и ландшафты бассейна Дона: реликтовые петрофитные сообщества Донской гряды и Донецкого кряжа, псаммофитная растительность донских песчаных массивов, аридные засоленные территории Южных Ергеней, древних долин Западного Маныча и Сала.
В результате геоботанического обследования Ростовской области были описаны 1116 геоботанических площадок (139 ключевых участков) по общепринятым методикам [10]. Для каждой площадки указывались географические координаты с использованием GPS и заносились в базу данных программы IBIS [11], в которой проводилась обработка валовых таблиц геоботанических описаний.
В результате синтаксономической проработки на основе подхода Браун-Бланке была выполнена эколого-флористическая классификация степной растительности бассейна Дона.
Сопоставления растительных единиц различных классификационных построений позволили обосновать филоценогенетические закономерности их развития и выявить фитоценотическое разнообразие степей в пределах Ростовской области [12].
Проведенные исследования позволили выделить в растительном покрове причерноморских и заволжско-казахстанских степей бассейна Дона 37 ассоциаций в составе 6 союзов, 4 порядков и 4 классов эколого-флористической классификации [12, 13].
С позиций эколого-флористической классификации сообщества настоящих разнотравно-дерновиннозлаковых, дерновиннозлаковых и опустыненных полукустарничково-дерновиннозлаковых причерноморских и заволжско-казахстанских степей [8, 9] отнесены к классу Festuco-Brometea Br.-Bl. et Tx. 1943; петрофитная растительность выделена в синтаксоны класса Helianthemo-Thymetea Romashchenko, Didukh et Solomakha 1996; псаммофитные сообщества выделенных синтаксонов рассматриваются в составе класса Festuceteavaginatae Soo em. Vicherek 1972; галофитная растительность солонцов и гемигалофитные дериватные сообщества представляют синтаксоны из состава класса Festuco-Puccinellietеa Soό ex Vicherek 1973.
Как видно из таблицы 1, в классе Festuco-Brometea были объединены все сообщества степного типа растительности. Всего в составе класса среди высших синтаксономических единиц выделяются 1 порядок, два союза и 4 подсоюза, однако ранг синтаксонов на уровне союзов в дальнейшем может быть пересмотрен на уровне порядков.
Таблица 1. Синтаксоны класса Festuco-Brometea
№ ассоциаций* |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
13 |
14 |
15 |
16 |
17 |
|
Количество описаний |
67 |
21 |
39 |
23 |
72 |
27 |
17 |
23 |
26 |
26 |
32 |
11 |
16 |
64 |
35 |
24 |
34 |
|
Диагностические виды ассоциаций |
|||||||||||||||||
|
Stipa tirsa |
V |
I |
II |
III |
. |
+ |
. |
+ |
. |
IV |
. |
+ |
+ |
. |
. |
. |
. |
|
Trifolium alpestre |
V |
III |
+ |
+ |
+ |
. |
II |
. |
. |
I |
. |
. |
. |
. |
+ |
. |
. |
|
Stipa capillata |
III |
V |
III |
. |
II |
III |
IV |
III |
III |
II |
IV |
II |
II |
+ |
+ |
II |
II |
|
Rosa sp. |
I |
IV |
+ |
. |
+ |
+ |
II |
II |
I |
II |
+ |
. |
. |
. |
. |
. |
. |
|
Polygala comosa |
I |
III |
+ |
. |
. |
. |
. |
. |
I |
. |
. |
. |
. |
. |
. |
. |
. |
|
Artemisia marschalliana |
III |
III |
V |
II |
. |
II |
III |
. |
II |
I |
. |
. |
. |
+ |
. |
+ |
+ |
|
Helichrysum arenarium |
II |
II |
IV |
. |
+ |
I |
II |
. |
I |
I |
. |
+ |
+ |
. |
. |
. |
. |
|
Jurinea cyanoides |
I |
+ |
IV |
. |
. |
+ |
. |
. |
. |
+ |
. |
. |
. |
. |
. |
. |
. |
|
Astragalus varius |
+ |
+ |
IV |
+ |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
|
Gypsophila paniculata |
I |
I |
III |
I |
+ |
+ |
. |
. |
+ |
+ |
+ |
+ |
+ |
. |
. |
. |
+ |
|
Pulsatilla patens |
+ |
+ |
III |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
|
Pulsatilla pratensis |
+ |
I |
III |
. |
. |
. |
+ |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
|
Bellevalia sarmatica |
I |
I |
. |
V |
III |
I |
+ |
III |
III |
II |
II |
II |
II |
III |
IV |
II |
. |
|
Pedicularis physocalyx |
. |
. |
. |
V |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
|
Peucedanum ruthenicum |
I |
II |
II |
V |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
|
Salvia stepposa |
II |
II |
. |
V |
. |
. |
+ |
. |
. |
+ |
. |
+ |
I |
. |
. |
. |
. |
|
Achillea submillefolium |
I |
+ |
I |
V |
+ |
. |
. |
. |
. |
. |
+ |
+ |
. |
. |
. |
. |
. |
|
Xanthoselinum alsaticum |
+ |
I |
+ |
III |
. |
. |
. |
+ |
+ |
I |
. |
+ |
+ |
. |
. |
. |
. |
|
Stipa lessingiana |
+ |
II |
. |
. |
V |
III |
II |
IV |
V |
III |
V |
III |
IV |
III |
IV |
V |
IV |
|
Oxytropis pilosa |
+ |
. |
. |
. |
III |
+ |
. |
. |
II |
II |
III |
I |
I |
. |
. |
. |
. |
|
Viola ambigua |
I |
I |
. |
+ |
III |
I |
. |
II |
III |
I |
II |
+ |
. |
. |
. |
. |
. |
|
Cephalaria uralensis |
I |
. |
II |
. |
II |
IV |
II |
. |
+ |
I |
+ |
. |
. |
. |
. |
. |
. |
|
Thymus dimorphus |
. |
. |
. |
. |
. |
III |
+ |
+ |
I |
+ |
. |
. |
. |
. |
. |
. |
. |
|
Cleistogenes bulgarica |
. |
. |
. |
. |
+ |
III |
I |
+ |
I |
. |
. |
. |
. |
. |
. |
. |
. |
|
Linum czerniaevii |
. |
. |
. |
. |
I |
III |
+ |
. |
I |
. |
. |
. |
. |
. |
. |
. |
. |
|
Jurinea stoechadifolia |
. |
. |
. |
. |
. |
III |
. |
. |
+ |
. |
. |
. |
. |
. |
. |
. |
. |
|
Scleranthus annuus |
. |
. |
. |
. |
. |
. |
V |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
|
Berteroa incana |
I |
+ |
+ |
. |
+ |
I |
III |
. |
. |
+ |
. |
+ |
I |
. |
. |
. |
+ |
|
Stipa zalesskii |
+ |
. |
. |
. |
I |
II |
II |
V |
I |
II |
III |
. |
. |
+ |
II |
I |
+ |
|
Bothriochloa ischaemum |
. |
. |
. |
. |
+ |
+ |
+ |
II |
+ |
+ |
. |
. |
. |
. |
. |
. |
. |
|
Stipa pulcherrima |
. |
. |
. |
. |
. |
+ |
. |
V |
V |
V |
IV |
. |
. |
. |
. |
. |
. |
|
Centaurea orientalis |
I |
II |
+ |
. |
II |
II |
I |
+ |
V |
II |
I |
. |
. |
. |
. |
. |
. |
|
Inula aspera |
. |
. |
. |
. |
+ |
+ |
+ |
. |
III |
+ |
. |
. |
. |
. |
. |
. |
. |
|
Plantago urvillei |
IV |
IV |
+ |
V |
IV |
II |
III |
I |
III |
V |
III |
II |
III |
. |
. |
. |
. |
|
Centaurea adpressa |
II |
I |
II |
I |
+ |
+ |
+ |
I |
. |
III |
. |
I |
II |
. |
. |
. |
+ |
|
Stachys atherocalyx |
. |
. |
. |
. |
I |
+ |
I |
I |
I |
III |
I |
. |
. |
. |
. |
+ |
. |
|
Campanula macrostachya |
. |
. |
. |
. |
. |
. |
. |
. |
. |
II |
. |
. |
. |
. |
. |
. |
. |
|
Astragalus ponticus |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
IV |
. |
. |
. |
. |
. |
. |
|
Adonis wolgensis |
+ |
. |
. |
IV |
+ |
I |
. |
+ |
+ |
I |
IV |
+ |
. |
. |
. |
. |
. |
|
Carduus nutans |
. |
. |
. |
. |
I |
. |
. |
. |
. |
. |
III |
+ |
. |
. |
. |
+ |
. |
|
Calophaca wolgarica |
. |
. |
. |
. |
. |
. |
. |
II |
. |
. |
III |
. |
. |
. |
. |
. |
+ |
|
Verbascum ovalifolium |
. |
. |
. |
. |
I |
. |
. |
. |
I |
. |
II |
. |
. |
. |
. |
. |
. |
|
Festuca pseudovina |
. |
. |
. |
. |
+ |
I |
. |
+ |
. |
. |
+ |
V |
II |
+ |
III |
II |
III |
|
Ajuga orientalis |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
II |
III |
. |
. |
. |
. |
. |
|
Astragalus asper |
. |
. |
. |
. |
. |
. |
. |
+ |
. |
+ |
. |
IV |
V |
. |
. |
. |
. |
|
Astragalus longipetalus |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
I |
II |
. |
. |
. |
. |
|
Tragopogon dasyrhynchus |
I |
. |
I |
II |
+ |
+ |
. |
I |
. |
II |
II |
III |
IV |
II |
I |
I |
+ |
|
Agropyron pectinatum |
. |
. |
. |
. |
. |
+ |
+ |
II |
+ |
. |
III |
+ |
I |
IV |
+ |
II |
I |
|
Astragalus reduncus |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
III |
I |
I |
I |
|
Chenopodium album |
. |
. |
+ |
. |
+ |
. |
. |
. |
. |
. |
. |
. |
. |
III |
+ |
. |
+ |
|
Agropyron desertorum |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
+ |
IV |
I |
II |
|
Cerastium syvaschicum |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
I |
IV |
I |
. |
|
Amoria retusa |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
II |
IV |
+ |
. |
|
Vicia hirsuta |
. |
. |
. |
. |
. |
. |
+ |
. |
. |
. |
. |
. |
. |
. |
III |
. |
+ |
|
Cruciata pedemontana |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
III |
. |
. |
|
Stipa ucrainica |
. |
. |
. |
. |
. |
III |
II |
III |
II |
III |
I |
+ |
+ |
II |
III |
V |
I |
|
Artemisia lerchiana |
. |
. |
. |
. |
+ |
II |
. |
. |
. |
. |
. |
I |
II |
. |
. |
+ |
V |
|
Artemisia pauciflora |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
+ |
. |
+ |
II |
|
Colchicum laetum |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
II |
|
Ceratocephala testiculata |
. |
. |
. |
. |
. |
+ |
. |
. |
. |
. |
+ |
+ |
. |
+ |
. |
+ |
II |
|
Д. в. подсоюза Festuco rupicolae–Stipenion pennatae |
|||||||||||||||||
|
Stipa pennata |
V |
V |
IV |
IV |
+ |
+ |
. |
+ |
. |
I |
. |
+ |
+ |
. |
. |
+ |
. |
|
Festuca rupicola |
IV |
V |
V |
V |
I |
II |
I |
II |
II |
I |
+ |
II |
II |
. |
. |
. |
. |
|
Potentilla humifusa |
IV |
IV |
IV |
IV |
I |
III |
II |
. |
I |
+ |
. |
+ |
I |
. |
. |
+ |
. |
|
Stipa dasyphylla |
III |
III |
V |
V |
. |
I |
II |
. |
. |
II |
. |
. |
. |
. |
. |
. |
. |
|
Trinia multicaulis |
III |
II |
II |
IV |
+ |
+ |
. |
. |
. |
+ |
. |
II |
II |
+ |
+ |
+ |
. |
|
Knautia arvensis |
II |
+ |
+ |
II |
. |
. |
+ |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
|
Ferulago galbanifera |
I |
I |
III |
IV |
. |
+ |
+ |
. |
. |
I |
I |
+ |
+ |
. |
. |
. |
. |
|
Inula hirta |
I |
I |
II |
III |
. |
. |
. |
. |
. |
. |
. |
+ |
+ |
. |
. |
. |
+ |
|
Д. в. подсоюза Phlomenion pungentis |
|||||||||||||||||
|
Euphorbia stepposa |
+ |
II |
. |
. |
III |
III |
+ |
+ |
V |
II |
. |
I |
+ |
. |
. |
. |
. |
|
Orthanthella lutea |
I |
I |
I |
. |
+ |
II |
II |
+ |
I |
I |
. |
+ |
I |
. |
. |
. |
. |
|
Galium octonarium |
V |
IV |
I |
IV |
III |
II |
II |
IV |
V |
V |
V |
I |
+ |
+ |
. |
+ |
+ |
|
Seseli tortuosum |
IV |
IV |
IV |
III |
II |
III |
. |
I |
I |
III |
IV |
III |
IV |
+ |
. |
+ |
+ |
|
Stachys recta |
IV |
IV |
IV |
V |
IV |
III |
IV |
II |
IV |
III |
IV |
II |
II |
. |
. |
+ |
. |
|
Bromopsis riparia |
IV |
IV |
III |
II |
IV |
IV |
III |
III |
V |
V |
V |
II |
I |
. |
. |
+ |
+ |
|
Veronica jacquinii |
IV |
III |
II |
IV |
III |
II |
II |
II |
III |
IV |
III |
III |
III |
. |
. |
+ |
. |
|
Erysimum canescens |
I |
II |
II |
. |
III |
II |
IV |
III |
III |
II |
V |
II |
II |
. |
+ |
I |
II |
|
Eryngium campestre |
V |
V |
IV |
II |
IV |
IV |
V |
IV |
III |
IV |
V |
IV |
V |
II |
II |
IV |
II |
|
Salvia tesquicola |
III |
III |
II |
. |
IV |
II |
III |
III |
III |
IV |
V |
IV |
IV |
II |
I |
III |
+ |
|
Phlomis pungens |
I |
+ |
+ |
V |
IV |
III |
II |
III |
IV |
III |
V |
III |
IV |
II |
III |
III |
II |
|
Goniolimon tataricum |
+ |
II |
+ |
II |
I |
+ |
II |
I |
I |
II |
+ |
II |
II |
II |
I |
+ |
+ |
|
Nepeta parviflora |
+ |
+ |
. |
II |
IV |
I |
. |
I |
III |
I |
IV |
+ |
+ |
+ |
I |
+ |
. |
|
Linum austriacum |
. |
. |
. |
. |
IV |
II |
I |
IV |
IV |
II |
V |
I |
I |
II |
II |
II |
II |
|
Tanacetum millefolium |
. |
. |
. |
. |
+ |
II |
II |
I |
II |
+ |
I |
+ |
I |
+ |
. |
. |
+ |
|
Marrubium praecox |
. |
. |
. |
. |
IV |
II |
+ |
II |
V |
+ |
IV |
. |
. |
. |
. |
+ |
+ |
|
Д. в. союза Festucion valesiacae |
|||||||||||||||||
|
Salvia nutans |
II |
II |
+ |
. |
V |
IV |
IV |
I |
V |
II |
I |
+ |
+ |
. |
. |
. |
. |
|
Achillea stepposa |
V |
V |
II |
. |
V |
III |
III |
III |
IV |
V |
V |
IV |
III |
+ |
. |
I |
+ |
|
Thesium arvense |
III |
III |
II |
I |
II |
III |
II |
I |
+ |
II |
I |
III |
III |
. |
+ |
+ |
I |
|
Limonium platyphyllum |
I |
+ |
. |
II |
+ |
+ |
+ |
II |
+ |
III |
IV |
II |
III |
. |
. |
+ |
. |
|
Astragalus onobrychis |
+ |
+ |
+ |
. |
II |
II |
. |
I |
IV |
+ |
I |
+ |
+ |
. |
. |
+ |
. |
|
Astragalus ucrainicus |
. |
+ |
. |
. |
III |
III |
. |
II |
II |
. |
+ |
II |
II |
+ |
. |
+ |
+ |
|
Astragalus austriacus |
. |
. |
. |
. |
III |
I |
+ |
. |
IV |
+ |
V |
+ |
. |
. |
. |
. |
. |
|
Д. в. союза Fragario viridis-Trifolion montani |
|||||||||||||||||
|
Elytrigia repens |
V |
V |
III |
V |
IV |
II |
II |
V |
IV |
V |
IV |
IV |
III |
III |
IV |
III |
I |
|
Amoria montana |
V |
V |
II |
III |
. |
+ |
I |
. |
. |
I |
. |
. |
. |
. |
. |
+ |
. |
|
Fragaria viridis |
V |
V |
II |
+ |
+ |
+ |
+ |
. |
+ |
+ |
. |
. |
. |
. |
. |
. |
+ |
|
Filipendula vulgaris |
IV |
IV |
II |
V |
+ |
+ |
II |
+ |
+ |
II |
. |
. |
. |
. |
. |
. |
+ |
|
Cichorium intybus |
II |
III |
+ |
. |
I |
I |
. |
II |
I |
II |
+ |
II |
+ |
+ |
. |
+ |
+ |
|
Centaurea pseudomaculosa |
II |
III |
+ |
. |
I |
+ |
I |
+ |
+ |
+ |
. |
II |
II |
+ |
. |
. |
. |
|
Genista tinctoria |
II |
III |
+ |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
|
Д. в. союза Poo bulbosae–Caricion stenophyllae |
|||||||||||||||||
|
Poa bulbosa |
. |
+ |
III |
. |
I |
III |
II |
II |
I |
I |
+ |
+ |
+ |
V |
V |
V |
V |
|
Carex stenophylla |
+ |
+ |
. |
. |
+ |
. |
I |
+ |
. |
. |
. |
. |
. |
+ |
III |
II |
III |
|
Д. в. союза Tanaceto achilleifolii–Artemision santonicae |
|||||||||||||||||
|
Tanacetum achilleifolium |
. |
. |
. |
. |
+ |
+ |
. |
+ |
. |
. |
I |
I |
I |
V |
IV |
III |
IV |
|
Artemisia santonica |
. |
+ |
+ |
. |
. |
+ |
. |
+ |
. |
. |
. |
+ |
+ |
V |
IV |
II |
III |
|
Serratula erucifolia |
. |
+ |
. |
I |
+ |
+ |
+ |
I |
. |
II |
III |
II |
II |
V |
V |
II |
I |
|
Bromus squarrosus |
+ |
. |
I |
. |
I |
I |
II |
II |
+ |
. |
IV |
II |
II |
IV |
IV |
III |
V |
|
Leymus ramosus |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
I |
I |
II |
III |
II |
I |
|
Crepis tectorum |
. |
. |
. |
. |
I |
+ |
+ |
. |
. |
. |
. |
+ |
. |
III |
IV |
II |
II |
|
Phlomoides puberula |
. |
. |
. |
. |
+ |
. |
. |
. |
. |
. |
+ |
. |
. |
II |
III |
II |
+ |
|
Trifolium diffusum |
. |
. |
. |
. |
+ |
. |
. |
. |
. |
+ |
. |
. |
. |
II |
III |
+ |
+ |
|
Ranunculus oxyspermus |
. |
. |
. |
. |
. |
. |
. |
+ |
. |
. |
+ |
. |
. |
III |
I |
III |
II |
|
Tulipa gesneriana |
. |
. |
. |
. |
. |
+ |
. |
+ |
. |
. |
+ |
. |
. |
III |
III |
II |
II |
|
Prangos odontalgica |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
III |
III |
+ |
I |
|
Д. в. подсоюза Trifolio arvensis–Limonienion sareptani |
|||||||||||||||||
|
Trifolium arvense |
+ |
. |
+ |
. |
. |
. |
II |
. |
. |
+ |
. |
. |
. |
IV |
V |
III |
+ |
|
Limonium sareptanum |
. |
. |
. |
. |
. |
+ |
. |
+ |
. |
. |
. |
. |
. |
III |
IV |
II |
+ |
|
Ventenata dubia |
. |
. |
. |
. |
. |
. |
+ |
. |
. |
. |
. |
. |
. |
+ |
III |
+ |
. |
|
Vicia villosa |
. |
. |
. |
. |
. |
. |
. |
+ |
. |
+ |
. |
. |
. |
III |
IV |
I |
. |
|
Pastinaca clausii |
. |
. |
. |
. |
. |
. |
. |
. |
. |
+ |
. |
. |
. |
III |
III |
+ |
. |
|
Lepidium perfoliatum |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
II |
III |
II |
+ |
|
Д. в. порядка Festucetaliavalesiacae и класса Festuco-Brometea |
|||||||||||||||||
|
Koeleria cristata |
IV |
V |
IV |
+ |
IV |
V |
IV |
IV |
IV |
V |
V |
III |
V |
II |
III |
V |
IV |
|
Galium verum |
IV |
V |
IV |
V |
+ |
I |
IV |
II |
I |
III |
II |
IV |
IV |
+ |
III |
II |
+ |
|
Poa angustifolia |
IV |
V |
II |
V |
IV |
I |
I |
III |
III |
V |
V |
III |
II |
. |
+ |
I |
+ |
|
Euphorbia virgata |
IV |
IV |
III |
. |
I |
II |
II |
II |
+ |
III |
I |
I |
+ |
. |
+ |
I |
+ |
|
Falcaria vulgaris |
IV |
IV |
II |
IV |
IV |
II |
II |
IV |
IV |
V |
V |
IV |
V |
III |
III |
IV |
II |
|
Galatella villosa |
IV |
II |
III |
V |
III |
IV |
III |
IV |
IV |
IV |
V |
III |
V |
IV |
V |
II |
II |
|
Medicago romanica |
III |
IV |
II |
III |
V |
IV |
IV |
V |
V |
V |
IV |
IV |
V |
II |
+ |
III |
I |
|
Convolvulus arvensis |
III |
IV |
I |
II |
IV |
II |
IV |
IV |
III |
III |
III |
III |
III |
+ |
. |
II |
. |
|
Thymus marschallianus |
III |
III |
II |
IV |
III |
II |
IV |
IV |
V |
V |
V |
I |
I |
+ |
+ |
II |
+ |
|
Nonea rossica |
III |
I |
+ |
+ |
II |
III |
II |
+ |
II |
I |
+ |
II |
II |
. |
. |
+ |
. |
|
Artemisia austriaca |
II |
III |
III |
+ |
III |
II |
IV |
IV |
II |
IV |
IV |
IV |
IV |
IV |
V |
V |
V |
|
Festuca valesiaca |
II |
IV |
I |
III |
V |
V |
V |
IV |
IV |
V |
V |
IV |
IV |
IV |
IV |
IV |
V |
|
Galium humifusum |
II |
II |
. |
. |
II |
I |
I |
III |
+ |
II |
I |
III |
III |
+ |
+ |
II |
I |
|
Odontites vulgaris |
II |
I |
+ |
. |
III |
+ |
I |
IV |
II |
II |
+ |
II |
I |
+ |
. |
+ |
. |
|
Euphorbia seguieriana |
I |
I |
III |
. |
II |
IV |
III |
III |
II |
+ |
V |
I |
+ |
. |
+ |
IV |
III |
|
Phlomoides tuberosa |
I |
I |
+ |
IV |
+ |
I |
I |
I |
I |
III |
II |
I |
+ |
+ |
. |
. |
. |
|
Linaria maeotica |
I |
+ |
+ |
. |
II |
I |
+ |
III |
+ |
II |
I |
III |
III |
. |
+ |
II |
I |
Примечание. Приводится по О.Н. Деминой [12]; *Ассоциации: 1. Trifolio alpestris–Stipetum tirsae, 2. Stipetum capillatae, 3. Artemisio marschallianae–Stipetum dasyphyllae, 4. Bellevalio sarmaticae–Stipetum pennatae, 5. Stipetum lessingianae, 6. Cephalario uralensidis-Thymetum dimorphi, 7. Sclerantho annui-Stipetum capillatae, 8. Medicago romanicae–Stipetum zalesskii, 9. Plantagini stepposae–Stipetum pulcherrimae, 10. Plantagini urvillei–Stipetum tirsae, 11. Astragalo ponticae–Brometum squarrosi, 12. Ajugo orientalis–Festucetum pseudovinae, 13. Astragalo asperi–Stipetum lessingianae, 14. Agropyrini pectinati–Poetum bulbosae, 15. Amorio retusae–Cerastietum syvaschici, 16. Eryngio campestris–Stipetum ucrainicae, 17. Artemisio lerchianae–Stipetum lessingianae
Новый союз Tanacetoachilleifolii–Artemisionsantonicae на данном уровне синтаксономического анализа рассматривается нами в составе порядка Festucetaliavalesiacaeи представляет заволжско-казахстанские опустыненные степи на слабозасоленных почвах Южных Ергеней и древней долины Западного Маныча. С одной стороны, по диагностической комбинации видов данный союз близок к союзу Poobulbosae–Caricionstenophyllae Saitov 1989; с другой – территориально и в синтаксономическом пространстве, граничит с союзом Agropyrion pectinati V.Golub et Uzhametskaja 1991.
В пределах исследуемой территории различаются три литологических варианта петрофитных, или тимьянниковых степей (Steppae petrophile): кальцепетрофитный, псаммопетрофитный и ксеропетрофитный.
Тимьянниковые степи Приазовья и Донецкого кряжа рассматриваются в составе подсоюзаPhlomenionpungentis (табл. 1, асс. 6, 7, 8).
Синтаксоны кальцефитных степных сообществ отрогов Средне-Русской возвышенности (Донской гряды и Калачской возвышенности) выделены в подсоюзе Bupleurofalcati–Gypsophilenionaltissimae, входящего в состав союза Festucionvalesiacae (табл. 2, асс. 1 и 2).
Петрофитные кальцефитные степи Приазовья и отрогов Донецкого кряжа на данном уровне синтаксономического анализа сопоставляются с новым, предварительно рассматриваемым подсоюзомCleistogenobulgaricae–Jurinenionstoechadifoliae, который представляет юринейниково-ковылково-ковыльные полукустарничково-кустарничково-дерновиннозлаковые сообщества (табл. 2, асс. 3, 4). Судя по диагностической комбинации видов, с одной стороны эти союзы граничит с синтаксонами союза Euphorbiocretophilae-Thymioncretacei, входящего в состав класса Helianthemo-Thymetea; с другой – с подсоюзом Phlomenion pungentis, из состава класса Festuco-Brometea.Эти сообщества отличаются выраженной кальцефитно-петрофитной природой, оригинальностью видового состава, часто имеют галофитный и дериватный характер и могут рассматриваться как отдельный подтип (Steppae creta-petrophile).Кальце-петрофитные сообщества степного типа выделенных синтаксонов развиваются на скелетных почвах, формирующихся на третичных рыхлых и более древних плотных известняках и мергелях, и рассматриваются как гемипетрофитные степи.
В отличие от них, синтаксоны класса Helianthemo-Thymetea представляют азональную петрофитную растительность (Petrophyton) обнажений пород различной литологии, или тимьянников (табл. 2, асс. 5 – 11), но имеют также зонально-азональную природу своего формирования.
Таблица 2. Петрофитные сообщества тимьянниковых степей и тимьянников
Ассоциации * |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
|
Количество описаний |
18 |
6 |
27 |
13 |
11 |
7 |
11 |
4 |
5 |
10 |
8 |
|
Диагностические виды ассоциаций класса Festuco-Brometea |
|||||||||||
|
Astragalus albicaulis |
V |
. |
. |
. |
. |
. |
II |
. |
II |
. |
V |
|
Agropyron pectinatum |
IV |
. |
+ |
. |
. |
. |
I |
. |
I |
. |
. |
|
Krascheninnikovia ceratoides |
III |
. |
. |
. |
. |
. |
+ |
. |
. |
. |
. |
|
Echinops ruthenicus |
II |
. |
. |
. |
. |
. |
+ |
. |
. |
. |
. |
|
Elytrigia trichophora |
. |
V |
. |
. |
. |
. |
. |
. |
. |
+ |
. |
|
Aster amellus |
I |
IV |
. |
II |
. |
. |
. |
. |
. |
+ |
. |
|
Verbascum marschallianum |
I |
V |
+ |
. |
. |
. |
. |
. |
. |
. |
. |
|
Origanum vulgare |
. |
III |
+ |
. |
. |
. |
. |
. |
. |
+ |
. |
|
Anemone sylvestris |
. |
II |
. |
. |
. |
. |
. |
. |
. |
+ |
. |
|
Thymus dimorphus |
. |
. |
V |
II |
. |
. |
. |
. |
. |
. |
. |
|
Artemisia lerchiana |
II |
. |
III |
. |
. |
. |
I |
. |
I |
I |
I |
|
Gypsophila glomerata |
. |
. |
III |
I |
. |
. |
. |
. |
. |
. |
. |
|
Convolvulus lineatus |
I |
. |
I |
V |
. |
. |
. |
. |
. |
. |
II |
|
Vincetoxicum maeoticum |
. |
. |
+ |
V |
. |
. |
. |
. |
. |
. |
. |
|
Caragana scythica |
. |
. |
+ |
II |
. |
. |
. |
. |
. |
. |
. |
|
Asperula montana |
. |
. |
+ |
IV |
. |
. |
. |
. |
. |
. |
. |
|
Poterium polygamum |
. |
. |
. |
II |
. |
. |
. |
. |
. |
. |
. |
|
Д. в. подсоюза Bupleuro falcati–Gypsophilenion altissimae |
|||||||||||
|
Gypsophila altissima |
IV |
IV |
+ |
II |
V |
. |
III |
V |
V |
II |
V |
|
Salvia nutans |
IV |
V |
V |
V |
IV |
II |
. |
. |
. |
. |
. |
|
Medicago lupulina |
III |
V |
III |
IV |
. |
II |
. |
. |
. |
I |
. |
|
Agrimonia eupatoria |
II |
V |
I |
II |
. |
. |
+ |
. |
. |
I |
. |
|
Centaurea pseudomaculosa |
II |
II |
. |
. |
. |
. |
. |
. |
. |
I |
. |
|
Jurinea arachnoidea |
II |
. |
III |
. |
. |
. |
. |
. |
. |
. |
. |
|
Polygala sibirica |
I |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
|
Д. в. подсоюза Cleistogeno bulgaricae–Jurinenion stoechadifoliae |
|||||||||||
|
Jurinea stoechadifolia |
. |
. |
V |
V |
IV |
. |
. |
. |
. |
. |
. |
|
Cleistogenes bulgarica |
. |
. |
IV |
IV |
. |
. |
. |
. |
. |
. |
I |
|
Galatella villosa |
III |
. |
IV |
IV |
I |
. |
+ |
. |
. |
. |
I |
|
Meniocus linifolius |
+ |
. |
III |
III |
II |
. |
. |
. |
. |
. |
. |
|
Hyacinthella pallasiana |
. |
. |
IV |
V |
III |
. |
. |
. |
. |
. |
. |
|
Linum tenuifolium |
. |
. |
II |
IV |
V |
. |
. |
. |
. |
. |
I |
|
Linum czerniaevii |
. |
. |
V |
V |
III |
. |
. |
. |
. |
. |
. |
|
Microthlaspi perfoliatum |
. |
. |
II |
III |
III |
. |
. |
. |
. |
. |
. |
|
Alyssum calycinum |
. |
. |
I |
II |
. |
II |
. |
. |
. |
. |
. |
|
Dianthus pseudarmeria |
. |
. |
II |
III |
. |
. |
. |
. |
. |
. |
. |
|
Scorzonera mollis |
. |
. |
IV |
I |
. |
. |
. |
. |
. |
+ |
. |
|
Д. в. союза Festucion valesiacae |
|||||||||||
|
Thesium arvense |
III |
V |
III |
V |
I |
IV |
+ |
V |
I |
II |
II |
|
Astragalus onobrychis |
II |
IV |
II |
+ |
. |
IV |
. |
. |
. |
+ |
. |
|
Astragalus austriacus |
. |
. |
II |
I |
. |
. |
. |
. |
. |
. |
. |
|
Д. в. порядка Festucetalia valesiacae и класса Festuco-Brometea |
|||||||||||
|
Festuca valesiaca |
II |
III |
V |
V |
I |
V |
. |
III |
I |
. |
. |
|
Artemisia austriaca |
III |
. |
I |
III |
I |
IV |
. |
. |
. |
. |
. |
|
Koeleria cristata |
II |
IV |
V |
IV |
. |
V |
. |
II |
. |
. |
. |
|
Euphorbia seguieriana |
III |
I |
V |
III |
I |
. |
+ |
. |
I |
+ |
V |
|
Campanula sibirica |
II |
IV |
III |
. |
III |
. |
. |
II |
. |
+ |
. |
|
Plantago urvillei |
II |
IV |
II |
II |
I |
. |
. |
. |
. |
. |
. |
|
Stipa capillata |
V |
III |
IV |
V |
II |
. |
. |
. |
I |
+ |
II |
|
Festuca rupicola |
II |
V |
II |
III |
. |
. |
. |
. |
. |
+ |
. |
|
Convolvulus arvensis |
+ |
III |
I |
I |
. |
V |
. |
. |
. |
+ |
. |
|
Galium verum |
III |
II |
. |
. |
. |
. |
+ |
. |
. |
I |
. |
|
Medicago romanica |
. |
. |
+ |
. |
I |
V |
. |
. |
I |
II |
. |
|
Asperula cynanchica |
+ |
. |
+ |
II |
. |
IV |
. |
. |
. |
. |
. |
|
Диагностические виды ассоциаций класса Helianthemo-Thymetea |
|||||||||||
|
Artemisia salsoloides |
II |
. |
. |
. |
V |
. |
III |
V |
V |
. |
IV |
|
Genista scythica |
. |
. |
II |
IV |
V |
. |
. |
. |
. |
. |
. |
|
Reseda lutea |
I |
. |
II |
II |
V |
II |
+ |
. |
. |
II |
I |
|
Onosma tanaitica |
III |
I |
+ |
. |
V |
. |
+ |
. |
. |
. |
II |
|
Alyssum tortuosum |
. |
. |
III |
. |
V |
V |
. |
. |
. |
. |
. |
|
Leontodon biscutellifolius |
. |
. |
I |
I |
IV |
. |
. |
. |
. |
. |
. |
|
Hedysarum grandiflorum |
. |
. |
I |
+ |
IV |
. |
. |
. |
. |
. |
. |
|
Centaurea ruthenica |
. |
. |
. |
. |
IV |
. |
. |
. |
. |
. |
. |
|
Erucastrum armoracioides |
. |
. |
. |
. |
III |
. |
. |
. |
. |
. |
. |
|
Linum hirsutum |
+ |
. |
+ |
. |
II |
. |
. |
. |
. |
. |
. |
|
Diplotaxis tenuifolia |
+ |
. |
. |
. |
II |
. |
. |
. |
. |
. |
. |
|
Hyssopus officinalis |
. |
. |
. |
. |
. |
V |
. |
. |
. |
. |
. |
|
Silene borysthenica |
. |
. |
. |
. |
. |
V |
. |
. |
. |
. |
. |
|
Bromus squarrosus |
I |
I |
I |
II |
. |
V |
. |
. |
I |
. |
. |
|
Achillea leptophylla |
. |
. |
I |
. |
. |
V |
. |
. |
. |
. |
. |
|
Elytrigia stipifolia |
. |
. |
. |
. |
. |
IV |
. |
. |
. |
. |
. |
|
Alyssum turkestanicum |
. |
. |
III |
. |
. |
IV |
. |
. |
. |
. |
. |
|
Xeranthemum annuum |
. |
. |
I |
. |
. |
IV |
. |
. |
. |
. |
. |
|
Anisantha tectorum |
. |
. |
. |
. |
. |
IV |
. |
. |
. |
. |
. |
|
Salvia verticillata |
. |
. |
+ |
I |
. |
IV |
. |
. |
. |
. |
. |
|
Echium vulgare |
. |
. |
+ |
. |
. |
IV |
. |
. |
. |
+ |
. |
|
Festuca pratensis |
. |
. |
. |
. |
. |
IV |
. |
. |
. |
+ |
. |
|
Matthiola fragrans |
+ |
I |
+ |
. |
. |
. |
V |
. |
. |
. |
III |
|
Asperula tephrocarpa |
. |
. |
II |
. |
IV |
. |
V |
. |
. |
+ |
IV |
|
Melilotus officinalis |
II |
IV |
. |
. |
. |
. |
V |
. |
. |
IV |
. |
|
Atraphaxis frutescens |
. |
. |
. |
. |
. |
. |
III |
. |
. |
. |
. |
|
Silene cretacea |
. |
. |
. |
. |
. |
. |
II |
. |
. |
. |
. |
|
Hedysarum cretaceum |
. |
. |
. |
. |
. |
. |
. |
V |
. |
. |
. |
|
Melica transsilvanica |
. |
I |
. |
. |
I |
. |
+ |
V |
. |
. |
. |
|
Artemisia absinthium |
. |
. |
. |
. |
. |
. |
. |
V |
. |
. |
. |
|
Poa angustifolia |
I |
IV |
+ |
+ |
. |
. |
I |
V |
. |
. |
. |
|
Poa compressa |
I |
II |
II |
. |
. |
III |
. |
V |
II |
III |
. |
|
Lepidium meyeri |
. |
. |
. |
. |
. |
. |
. |
. |
V |
. |
. |
|
Scrophularia cretacea |
. |
. |
. |
. |
. |
. |
. |
III |
V |
II |
II |
|
Festuca cretacea |
. |
. |
. |
. |
. |
. |
I |
. |
II |
V |
. |
|
Erysimum cretaceum |
. |
. |
. |
. |
. |
. |
. |
. |
. |
II |
. |
|
Artemisia hololeuca |
. |
. |
. |
. |
. |
. |
. |
. |
. |
. |
V |
|
Linum ucranicum |
I |
. |
. |
. |
. |
. |
. |
. |
. |
. |
V |
|
Plantago salsa |
+ |
. |
. |
. |
. |
. |
II |
. |
. |
+ |
IV |
|
Silene supina |
+ |
. |
. |
IV |
. |
. |
II |
. |
. |
. |
IV |
|
Orthanthella lutea |
I |
II |
III |
. |
I |
. |
. |
. |
. |
II |
IV |
|
Д.в. союза Euphorbio cretophilae-Thymion cretacei |
|||||||||||
|
Thymus calcareus |
II |
. |
. |
V |
V |
V |
II |
. |
. |
. |
V |
|
Euphorbia cretophila |
. |
. |
III |
V |
V |
. |
. |
. |
. |
. |
. |
|
Erucastrum cretaceum |
. |
. |
. |
. |
I |
. |
. |
. |
. |
+ |
. |
|
Д.в. союза Centaureo carbonatae-Koelerion talievii |
|||||||||||
|
Centaurea carbonata |
III |
. |
. |
V |
. |
. |
. |
. |
I |
. |
I |
|
Koeleria talievii |
. |
. |
. |
. |
. |
. |
+ |
III |
I |
+ |
. |
|
Teucrium polium |
III |
I |
V |
V |
V |
V |
. |
. |
. |
. |
. |
|
Д.в. союза Artemisio hololeucae-Hyssopion cretacei |
|||||||||||
|
Polygala cretacea |
I |
I |
+ |
. |
II |
. |
. |
. |
. |
II |
I |
|
Д.в. класса Helianthemo-Thymetea и порядка Thymo cretacei-Hissopetalia cretacei |
|||||||||||
|
Pimpinella tragium |
II |
I |
IV |
V |
V |
V |
V |
V |
V |
V |
V |
|
Cephalaria uralensis |
III |
. |
V |
V |
IV |
IV |
IV |
III |
IV |
+ |
V |
|
Hyssopus cretaceus |
. |
. |
. |
. |
. |
. |
V |
V |
V |
III |
V |
|
Bupleurum falcatum |
+ |
. |
. |
. |
. |
. |
II |
. |
. |
+ |
. |
Примечание. Приводится по О.Н. Деминой [12]; Ассоциации*: 1 – Astragalo albicaulis–Stipetum capillatae; 2 – Astro amelli–Elytrigetum trichophorae; 3 – Cephalario uralensidis–Thymetum dimorphi; 4 – Convolvulis lineati–Vincetoxietum maeotici; 5 – Genisto scythicae–Artemisietum salsoloidis; 6 – Sileno borysthenicae–Hyssopetum angustifolii; 7 – Matthiolo fragransi–Atraphaxietum frutescens; 8 – Hedysaro cretacei–Melicetum transsilvanicae; 9 – Lepidio meyeri–Scrophularietum cretacei; 10 – Erysimo cretacei–Festucetum cretacei; 11 – Artemisio hololeucae–Polygaletum cretaceae.
Сообщества псаммофитона (Psammophyton) на песчаных массивах в долинах Дона и его притоков, в отличие от гемипсаммофитных сообществ степного типа (Steppae ammophile), представляют азональную растительность и отнесены к синтаксонам, входящим в состав класса Festuceteavaginatae (табл. 3). Фактор увлажнения и сукцессионный статус сообществ играют ведущую роль в дифференциации ценозов на песках, что обусловливает их синтаксономическое разнообразие.
Таблица 3. Псаммофитные сообщества синтаксонов класса Festucetea vaginatae
|
Ассоциации * |
1 |
2 |
3 |
4 |
5 |
6 |
|
Количество описаний |
24 |
42 |
12 |
33 |
38 |
5 |
|
Диагностические виды ассоциаций |
||||||
|
Potentilla arenaria |
IV |
I |
II |
I |
. |
. |
|
Hieracium echioides |
IV |
II |
I |
I |
I |
. |
|
Scirpoides holoschoenus |
+ |
V |
II |
I |
+ |
. |
|
Calamagrostis epigeios |
III |
V |
I |
II |
II |
. |
|
Potentilla argentea |
III |
V |
+ |
I |
. |
. |
|
Galium verum |
II |
V |
II |
+ |
. |
I |
|
Myosotis micrantha |
II |
V |
II |
+ |
+ |
I |
|
Verbascum phoeniceum |
I |
V |
. |
+ |
. |
. |
|
Genista sibirica |
. |
V |
. |
+ |
+ |
. |
|
Eryngium planum |
. |
IV |
. |
. |
. |
. |
|
Silene chlorantha |
I |
IV |
I |
. |
. |
. |
|
Asparagus officinalis |
+ |
IV |
. |
+ |
. |
. |
|
Hypericum perforatum |
+ |
IV |
. |
+ |
+ |
. |
|
Chondrilla graminea |
II |
II |
V |
III |
I |
. |
|
Anisantha tectorum |
I |
. |
V |
+ |
+ |
I |
|
Agropyron fragile |
+ |
I |
IV |
II |
+ |
. |
|
Syrenia montana |
II |
. |
IV |
I |
+ |
I |
|
Plantago arenaria |
. |
. |
III |
. |
. |
. |
|
Xanthium californicum |
. |
. |
III |
. |
. |
I |
|
Anthemis ruthenica |
+ |
. |
III |
+ |
. |
I |
|
Astragalus borysthenicus |
. |
. |
II |
. |
. |
. |
|
Centaurea gerberi |
+ |
. |
I |
V |
. |
. |
|
Agropyron tanaiticum |
I |
I |
II |
V |
I |
. |
|
Rumex acetosella |
III |
+ |
+ |
V |
I |
. |
|
Salix rosmarinifolia |
. |
I |
+ |
IV |
+ |
. |
|
Dianthus squarrosus |
. |
. |
. |
+ |
III |
. |
|
Linaria dulcis |
. |
. |
. |
II |
III |
. |
|
Cleistogenes squarrosa |
II |
. |
II |
. |
. |
V |
|
Allium atroviolaceum |
. |
. |
. |
. |
. |
V |
|
Scorzonera mollis |
. |
. |
. |
. |
. |
V |
|
Potentilla astracanica |
. |
+ |
. |
. |
. |
V |
|
Iris pumila |
+ |
. |
. |
. |
. |
IV |
|
Alyssum turkestanicum |
+ |
. |
I |
II |
+ |
V |
|
Bromus squarrosus |
I |
+ |
+ |
+ |
. |
IV |
|
Carex praecox |
. |
. |
. |
. |
. |
IV |
|
Filago arvensis |
. |
. |
. |
+ |
. |
IV |
|
Д. в. подсоюза Chamaecytiso borysthenici–Artemisienion arenariae |
|
|||||
|
Artemisia arenaria |
+ |
II |
+ |
V |
V |
V |
|
Tragopogon tanaiticus |
I |
+ |
+ |
V |
IV |
. |
|
Leymus racemosus |
+ |
. |
+ |
II |
III |
. |
|
Asperula graveolens |
+ |
. |
I |
III |
III |
. |
|
Chamaecytisus borysthenicus |
. |
. |
I |
IV |
II |
. |
|
Д.в. союза Festucion beckeri |
|
|
|
|
|
|
|
Festuca beckeri |
IV |
+ |
IV |
V |
V |
V |
|
Thymus pallasianus |
III |
II |
IV |
IV |
III |
V |
|
Koeleria sabuletorum |
V |
I |
IV |
V |
II |
III |
|
Achillea micrantha |
IV |
I |
V |
IV |
II |
I |
|
Jurinea cyanoides |
IV |
+ |
V |
II |
II |
III |
|
Seseli tortuosum |
IV |
V |
I |
II |
+ |
. |
|
Linaria genistifolia |
III |
III |
I |
+ |
. |
IV |
|
Carex colchica |
III |
III |
III |
V |
IV |
. |
|
Scabiosa ucranica |
II |
V |
I |
I |
+ |
. |
|
Senecio borysthenicus |
III |
IV |
+ |
I |
+ |
. |
|
Scorzonera ensifolia |
II |
+ |
II |
+ |
. |
. |
|
Agropyron lavrenkoanum |
II |
+ |
+ |
. |
. |
. |
|
Д.в. порядка Festucetalia vaginatae |
|
|
|
|
|
|
|
Secale sylvestre |
I |
III |
IV |
IV |
I |
. |
|
Astragalus varius |
IV |
II |
IV |
. |
+ |
V |
|
Anchusa popovii |
II |
. |
+ |
+ |
. |
. |
|
Taeniopetalum arenarium |
I |
. |
. |
. |
. |
. |
|
Д.в. класса Festucetea vaginatae |
|
|
|
|
|
|
|
Artemisia marschalliana |
V |
V |
V |
II |
+ |
I |
|
Stipa borysthenica |
V |
III |
V |
I |
I |
III |
|
Euphorbia seguieriana |
V |
III |
II |
V |
V |
III |
|
Helichrysum arenarium |
V |
II |
V |
V |
III |
V |
|
Silene borysthenica |
III |
. |
I |
II |
+ |
II |
|
Gypsophila paniculata |
II |
I |
V |
II |
II |
IV |
|
Chondrilla juncea |
I |
. |
. |
. |
. |
. |
Примечание. Приводится по О.Н. Деминой [12]; Ассоциации*: 1 – Hieracio echioidis–Stipetum borysthenicae; 2 – Scirpoido holoschoenii–Genistaetum sibiricae; 3 – Secalo–Stipetum borysthenicae; 4 – Centaureo gerberi–Agropyretum tanaiticae; 5 – Artemisio arenariae–Dianthetum squarrosi; 6 – Artemisio arenariae–Potentilletum astracanicae.
Синтаксоны класса Festuco-Puccinellietea Soό ex Vicherek 1973 –это азональная галофитная растительность на солонцах, которая рассматривается как часть галофитона (Halophyton, Salineta).
Для достижения поставленной цели были использованы методы непрямой ординации. В нашу задачу входило выявление главных факторов, влияющих на распределение сообществ степной растительности бассейна Дона и определение координации выделенных синтаксонов в пространстве экологических факторов [12]. Ординационный градиентный анализ данных был проведен с использованием непрямой ординации по методу DСA (detrended correspondence analysis) – бестрендовый анализ соответствия или метод взаимного усреднения [14], и NMDS (nonmetrical multidimensional scaling) – неметрическое многомерное шкалирование [15], вошедших в пакеты Juice 7.0.42 и PC-ORD [16], с предварительной обработкой в TWINSPAN [17].
Пространственные особенности растительного покрова исследовались с использованием ординационного анализа на основе выполненной синтаксономии степной растительности с применением эколого-флористических критериев [12] и построения картографической модели растительности степей бассейна Дона.
Результаты ординации
По результатам ординации 1116 описаний степной растительности бассейна Дона в пределах Ростовской области (рис. 1) по методу DСA (бестрендовый анализ соответствия) в пакете программ Juice 7.0.42, мера значимости (eigenvalue) первой оси ординации (DCA 1) оказалась равной 0,53; второй (DCA 2) – 0,52; третьей (DCA 3) – 0,23, четвертой – 0,20. По результатам ординации по методу DСA, выполненной в PC-ORD (рис. 2), мера значимости (eigenvalue) первой оси ординации (Axis 1) равняется 0,72; второй (Axis 2) – 0,64; третьей (Axis 3) – 0,41. Таким образом, только первые две оси ординации (рис. 1, 2) оказались экологически значимыми и могут отражать экологически верную информацию [18].
Известно, что методы непрямой ординации особенно эффективны в том случае, когда удается однозначно интерпретировать выделенные оси по составу концевых групп видов или сообществ [5]. В связи с этим, оси ординации были интерпретированы по составу концевых групп сообществ (Q-ординация) как оси факторальных нагрузок, или оси максимального варьирования (оси вариаций) в экологическом пространстве, отражающие комплексные градиенты факторов среды. Наибольшую нагрузку, как видно из рисунков (рис. 1, 2) и выше приведенных мер значимости, имеет первая ось (DCA 1, Axis 1) и представляет собой комплексный градиент бедности – богатства почв по степени увеличения кальцефильности.

Рис. 1 Положение 1116 описаний в осях ординации
(по методу DСA, Juice 7.0.42)
Примечание. Красные цифры – номера описаний; черные – номера фитоценонов.
На левом крае первой оси расположились псаммофитные сообщества, развивающиеся на песках, представленные псаммофитной растительностью песчаных массивов бассейна Дона (фитоценоны 1 – 20, рис. 1). То есть эти сообщества представляют концевую группу описаний на левом крае первой оси, где сгруппировались сообщества синтаксонов (класс II, рис. 2) в ранге ассоциаций: 1. Artemisio arenariae–Festucetum beckeri; 2. Centaureogerberi-Agropyretumtanaitici; 3. Hieracioechioidis–Stipetumborysthenicae; 4. Scirpoido–Genistaetum sibiricae; 5. Secalo–Stipetum borysthenicae; 6. Artemisio arenariae–Potentilletum astracanicae, входящих в состав класса Festucetea vaginatae.
На крайней правой позиции концевую группу описаний в экологическом пространстве с нагрузкой на первую ось представляют кальцефильные сообщества петрофитной растительности на мелах (фитоценоны 26 – 31, рис. 1). Это выделенные сообщества ассоциаций: 19. Matthiolofragrandis–Atraphaxietumfrutescens; 20. Hedysarocretacei–Melicetumtranssilvanicae; 21. Lepidio meyeri–Scrophularietum cretacei; 22. Artemisio hololeucae–Polygaletum cretaceae; 23. Centaureo carbonatae–Onosmetum tanaiticae; 24. Erysimo cretacei–Festucetum cretacei; 28. Genista scyticae–Artemisietum salsoloidis, входящие в состав класса Helianthemo-Thymetea (класс IV, рис. 2).
Таким образом, первая ось вариации представляет комплексный градиент смены псаммофитной растительности гемипсаммофитной, затем пелитофитной, несколько смещенной по центру и вытянутой дугой в сторону кальцефильности, и далее, на другом конце оси, петрофитной растительностью, по степени увеличения кальцефильности.
По центру первой оси (рис. 1, 2) расположились сообщества пелитофитных вариантов разнотравно-дерновиннозлаковых, дерновиннозлаковых и полукустарничково-дерновиннозлаковых степей, несущие нагрузку на вторую ось вариации (DCA 2, Axis 2).
Расположение концевых групп сообществ с нагрузкой на вторую ось вариации характеризует комплексный градиент общего увлажнения и богатства-засоленности почв (рис. 2). Внизу по оси (DСA 2, Axis 2) находится кластер наиболее мезофильных сообществ богаторазнотравно-дерновиннозлаковых восточноевропейских степей на обыкновенных и южных черноземах. Это многовидовые степные и лугово-степные сообщества из состава класса Festuco-Brometea, выделенные в ассоциации: 7. Trifolio alpestris–Stipetum tirsae; 8. Stipetum capillatae; 9. Artemisio marschallianae–Stipetum dasyphyllae; 10. Bellevaliosarmaticae–Stipetumpennatae.
Вверху с нагрузкой на вторую ось образуют концевую группу гемигалофитные и галофитные сообщества на солонцеватых и засоленных почвах (фитоценоны 41 – 47, рис. 1). Это степные дерновиннозлаковые и полукустарничково-дерновиннозлаковые сообщества ассоциаций: 33. Artemisiolerchianae–Stipetumlessingianae; 35. Amorioretusae–Cerastietumsyvaschici; 36. Agropyrinipectinati–Poetumbulbosae, входящие в состав класса Festuco-Brometea и галофитные маловидовые сообщества на солонцах ассоциации 37. Poobulbosae–Artemisietumpauciflorae из состава класса Festuco-Puccinellietea, занимающие самую крайнюю позицию в концевой галофитной группе.

Рис. 2. Положение 1116 геоботанических описаний в 1 и 2 осях ординации (по методу DСA, PC-ORD)
Примечание.Приводится по О.Н. Деминой [12]; Цифрами в тексте указаны номера ассоциаций: 1. Artemisio arenariae–Festucetum beckeri; 2. Centaureogerberi–Agropyretumtanaitici; 3. Hieracioechioidis–Stipetumborysthenicae; 4. Scirpoido–Genistaetum sibiricae; 5. Secalo–Stipetum borysthenicae; 6. Artemisio arenariae–Potentilletum astracanicae; 7. Trifolio alpestris–Stipetum tirsae; 8. Stipetum capillatae; 9. Artemisio marschallianae–Stipetum dasyphyllae;10. Bellevalio sarmaticae–Stipetum pennatae; 11. Plantagini urvillei–Stipetum tirsae; 12. Medicago romanicae–Stipetum zalesskii; 13. Plantagini stepposae–Stipetum pulcherrimae; 14. Astragalo ponticae–Brometum squarrosi; 15. Ajugo orientalae–Festucetum pseudovinae; 16. Astragalo asperi–Stipetum lessingianae; 17. Medicago romanicae–Stipetum ucrainicae; 18. Stipetum lessingianae; 19. Matthiolo fragrandis–Atraphaxietum frutescens; 20. Hedysaro cretacei–Melicetum transsilvanicae; 21. Lepidio meyeri–Scrophularietum cretacei; 22. Artemisio hololeucae–Polygaletum cretaceae; 23. Centaureo carbonatae–Onosmetum tanaiticae; 24. Erysimo cretacei–Festucetum cretacei; 25. Astragalo albicaulis–Stipetum capillatae; 26. Astro amelli–Elytrigetum trichophorae; 27. Medicago romanicae–Festucetum valesiacae; 28. Genista scyticae–Artemisietum salsoloidis; 29. Sileno borysthenicae–Hyssopetum angustifolii; 30. Cephalario uralensidis–Thymetum dimorphi; 31. Convolvulis lineati–Vincetoxietum maeotici; 32. Genisto scythicae–Stipetum adoxae; 33. Artemisio lerchianae–Stipetum lessingianae; 34. Eryngium campestre–Stipetum ucrainicae; 35. Amorio retusae–Cerastietum syvaschici; 36. Agropyrini pectinati–Poetum bulbosae; 37. Poobulbosae–Artemisietumpauciflorae.
Смещение значений второй оси символизирует градиент увеличения кальцефильности к югу, в богаторазнотравно- и разнотравно-дерновиннозлаковых сообществах, главным образом, западнопричерноморских степей. Обусловлено это тем, что они расположены не только южнее относительно первых, восточнопричерноморских, но и формируются на обыкновенных и южных черноземах, с близким залеганием мелов и известняков. Далее, по центру второй оси находятся разреженные кальцефильные петрофитные сообщества на мелах. Эти сообщества выделяются в синтаксонах из состава класса Helianthemo-Thymetea(класс IV, рис. 2) и представляют нагрузку на третью ось ординации. Концевая группа, расположенная по третьей оси внизу представляет петрофитные сообщества южной части области. Они формируются в бассейнах рек Северский Донец и Тузлов, на размытых мелах туронского яруса, сарматских известняках и выделяются в ассоциацию 22. Artemisio hololeucae–Polygaletum cretaceae и ассоциацию 28. Genista scyticae–Artemisietum salsoloidis.К этой группе тяготеют петрофитные сообщества ассоциации 29. Silenoborysthenicae–Hyssopetumangustifolii, которые развиваются в бассейне реки Тузлов и приурочены к каменноугольным песчано-глинистым сланцам южного макросклона Донецкого кряжа.
Вверху сгруппировались сообщества ассоциации 20. Hedysarocretacei–Melicetumtranssilvanicae, ассоциации 21. Lepidiomeyeri–Scrophularietumcretacei и ассоциации 24. Erysimocretacei–Festucetumcretacei, местообитания которых находятся на севере области, в бассейне Среднего Дона и связаныс мелами сеноманского яруса. Третья ось вариации рассматривается нами как комплексный долготный градиент, где важнейшим фактором выступает литолого-эдафический.
Как и следовало ожидать [18], ординация по методу DСA, которая была осуществлена в различных пакетах программного обеспечения, показала сходные результаты. Значения осей нагрузок оказались примерно одинаковыми как в пакете программы PC-ORD, так и Juice 7.0.42. Собственные величины осей ординации свыше 0,5 были определены только у первой и второй осей, которые оказались наиболее значимыми в экологическом пространстве. Однако определенное значение имеет и третья ось нагрузки (мера значимости в пакете программ PC-ORD имеет высокое значение – 0,41, хотя и ниже 0,5). В связи с этим, был применен NMDS – метод неметрического многомерного шкалирования, который иногда дает лучшие результаты ординации, чем DСA и является для последнего альтернативой. Он считается одним из наиболее оригинальных объективных методов, который рекомендуется использовать в работе с большим объемом материала [19, 20], как оказалось и в нашем случае, когда в исследовании экологии сообществ степной растительности бассейна Дона, при одновременной обработке использовалось 1116 геоботанических описаний. Значение стресса оказалось равным 14,89, что позволяет использовать данный метод ординационного анализа и означает небольшую разницу между модельной и эмпирической матрицами.

Рис. 3. Распределение 1116 описаний степной растительности в 1 и 2 осях ординации
(по методу NMDS, PC-ORD)
Примечание.Номера и названия ассоциаций приведены в примечании к рис. 2.
Непрямая ординация, то есть расположение сообществ ассоциаций в экологическом пространстве без прямой оценки экологических факторов, были также выполнены по методу NMDS, в программе PC-ORD (рис. 3). Полученные данные были использованы для интерпретации полученных результатов. Различия в вариабельности объектов (различия мер значимости осей, или значений нагрузок на заданные оси) показали, что пелитофитные варианты разнотравно-дерновиннозлаковых и дерновиннозлаковых степей в экологическом пространстве расположились в центре, в пределах меры значимости от -0,5 до +0,5.Далее, в трех заданных осях экологических факторов, в пределах меры значимости от -1 до +1, уже находятся степные гемипсаммофитные, гемипетрофитные и гемигалофитные сообщества синтаксонов, входящих в состав класса Festuco-Brometea.
Сообщества псаммофитной, петрофитной и галофитной растительности, имеющей зонально-азональную природу формирования и представляющей синтаксоны эколого-флористической классификации в ранге ассоциаций, входящие в состав классов Festuceteavaginatae,Helianthemo-Thymeteaи Festuco-Puccinellieteaсоответственно (рис. 3), расположились за пределами порогового значения данного показателя (от -1 до +1).
Проведенный ординационный анализ позволил:
- выявить три оси максимального варьирования в экологическом пространстве и интерпретировать эти оси вариации, которые отражают комплексные градиенты экологических факторов, влияющих на разнообразие степной растительности бассейна Дона;
- определить положение синтаксонов в ранге ассоциаций на комплексных градиентах среды, определяющих высокое разнообразие степной, псаммофитной, петрофитной и галофитной растительности, которое в целом формируется за счет общего увлажнения, богатства почв и засоления, литолого-эдафического фактора, в том числе степени кальцефитности почв и почвообразующих пород.
Построенная модель распределения степной растительности бассейна Дона в экологическом пространстве, созданная с применением ординационных подходов, отражает как непрерывность изменений растительного покрова на комплексных пространственных градиентах важнейших экологических факторов среды, так и разбиение его на отдельные, дискретные растительные единицы, согласно современным представлениям о единстве континуальности и дискретности растительного покрова [4, 21, 22, 23].
Пространственная модель растительного покрова
Построение экологической модели экотопической приуроченности и климатической обусловленности потребовало создания территориальных схем распределения растительности степей бассейна Дона (в пределах Ростовской области) на картографической основе, так как именно на карте моделируются пространственные особенности растительного покрова [24]. Построение картографической модели как особой географической формы моделирования, которая дополняет функциональную модель растительного сообщества и его ординацию в пространстве экологических факторов, осуществлялась вначале на основе растровых ГИС-моделей. Следующим этапом работ была «отмывка» растительности исследуемой территории на рельефной основе (рис. 4).
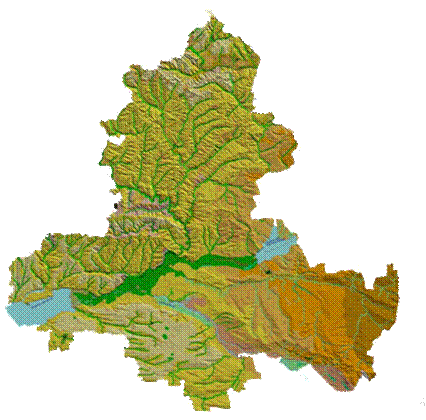
Рис. 4. ГИС-проект карты восстановленной растительности Ростовской области на рельефной основе

Рис. 5. Полигоны сохранившейся растительности Ростовской области в Google Планета Земля
При создании картографических моделей пространственной организации растительного покрова степей бассейна Дона использовались информационные технологии, при которых дешифрирование космоснимков выполнялось в Google Планета Земля (рис. 5). Затем полученные данные вносились в созданный ГИС-проект карты восстановленной растительности (рис. 4). Оцифровка геоботанических карт растительности сохранившихся участков степей и внедрение их в ГИС была проведена на последующем этапе работ.
ГИС-проект «Карта современной растительности Ростовской области» (рис. 6) создан в пакете программ ArcGIS 9.3 с использованием оригинальных первичных геоботанических данных, занесенных в программу IBIS, в среде которой была создана база данных (1116 геоботанических описаний с географическими координатами).
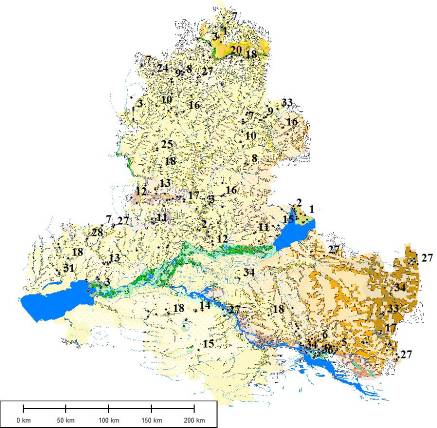
Рис. 6. Пространственное распределение степных сообществ выделенных ассоциаций в ГИС-проекте Карта растительности Ростовской области
Примечание. Цифрами обозначены номера ассоциаций, приведенные в примечании к рис. 2.
Построенная результирующая картографическая модель растительности отражает ее пространственную организацию, экотопическую приуроченность и климатическую обусловленность (рис. 6). Такой подход строился на основе интеграции слоев дешифрирования космической информации различных вегетационных периодов, цифровых моделей растительного покрова и рельефа.Все псаммофитные сообщества синтаксонов из состава класса Festuceteavaginatae, приуроченные к песчаным массивам бассейна Дона, оказались четко дифференцированы на территории Ростовской области и характеризуются преимущественно географической специфичностью и климатической обусловленностью (рис. 6). Ассоциации 1. Artemisioarenariae–Festucetumbeckeri и 3. Hieracioechioidis–Stipetumborysthenicae определяются для песчаных массивов Среднего Дона; ассоциации 2. Centaureogerberi–Agropyretumtanaitici и 4. Scirpoido–Genistaetumsibiricae – для песчаных массивов в долине Нижнего Дона.
Показано, что в отличие от них, сообщества ассоциации 6. Artemisioarenariae–Potentilletumastracanicae развиваются исключительно на юго-востоке, в подзоне опустыненных степей, то есть имеют отличия более высокого ранга, на подзональном уровне.
Пространственное распределение петрофитной растительности в ГИС-проекте Карта растительности Ростовской области характеризуется долготным и высотным градиентом. Сообщества петрофитной растительности, выделенные в ассоциацию 19. Matthiolofragrandis–Atraphaxietumfrutescens, ассоциацию 20. Hedysarocretacei–Melicetumtranssilvanicae, ассоциацию 21. Lepidiomeyeri–Scrophularietumcretacei, ассоциацию 22. Artemisiohololeucae–Polygaletumcretaceae, ассоциацию 23. Centaureocarbonatae–Onosmetumtanaiticae и ассоциацию 24. Erysimocretacei–Festucetumcretacei, из состава класса Helianthemo-Thymetea, а также гемипетрофитные сообщества ассоциации 25. Astragaloalbicaulis–Stipetumcapillatae из состава класса Festuco-Brometea, приурочены к наиболее повышенной северо-западной части области и развиваются на отрогах Средне-Русской возвышенности (Донская меловая гряда).
Сообщества петрофитной растительности, выделенные в ассоциации: 28. Genistascyticae–Artemisietumsalsoloidis; 29. Silenoborysthenicae–Hyssopetumangustifolii из класса Helianthemo-Thymetea и гемипетрофитные сообщества ассоциации 30. Cephalariouralensidis–Thymetumdimorphi, ассоциации 31. Convolvulislineati–Vincetoxietummaeotici и ассоциации 32. Genistoscythicae–Stipetumadoxae из класса Festuco-Brometea, отличаются относительно первых более южным своим расположением, в северо-восточном Приазовье и приурочены к южному макросклону Донецкого кряжа.
Сообщества ассоциации 7. Trifolioalpestris–Stipetumtirsae, ассоциации 8. Stipetumcapillatae, ассоциации 9. Artemisiomarschallianae–Stipetumdasyphyllae, ассоциации 10. Bellevaliosarmaticae–Stipetumpennatae и ассоциации 11. Plantaginiurvillei–Stipetumtirsae, представляющие гемипсаммофитные и пелитофитные варианты северных восточнопричерноморских степей (рис. 6),также хорошо дифференцированы, однако здесь отмечается их захождение по Донецкому кряжу в пределы распространения западнопричерноморских степей, расположенных южнее.
Закономерности пространственного распределения сообществ различных региональных типов восточнопричерноморских и западнопричерноморских степей в ГИС-проекте Карта растительности Ростовской области, кроме ботанико-географической специфики, также отражают особенности рельефа и их подзональной приуроченности. Сообщества ассоциации 16. Astragaloasperi–Stipetumlessingianae, котораяпредставляет восточнопричерноморские дерновиннозлаковые степи, иногда отмечаются в подзоне разнотравно-дерновиннозлаковых степей. Они развиваются в своеобразных реликтовых местообитаниях, связанных с выходами соленосных глин и распространены к северу от долины Нижнего Дона.
Сообщества западнопричерноморских степей ассоциации 13. Plantaginistepposae–Stipetumpulcherrimaeтяготеют к северо-восточному Приазовью и Донецкому кряжу; ассоциации 14. Astragaloponticae–Brometumsquarrosi, распространены исключительно к югу от долины Нижнего Дона.
Сообщества других ассоциаций: 12. Medicagoromanicae–Stipetumzalesskii; 17. Medicagoromanicae–Stipetumucrainicae; 18. Stipetumlessingianae, имеют климатическую обусловленность, широко распространены в подзоне разнотравно-дерновиннозлаковых степей и не обладают локальной ботанико-географической спецификой.
Пространственное распределение гемигалофитных и галофитных сообществ также обусловлено как климатическими факторами, так и большое значение в их развитии имеет экотопическая приуроченность (рис. 6). Эти степи представлены различными причерноморскими и заволжско-казахстанскими региональными ботанико-географическими типами.
Сообщества ассоциаций 33. Artemisio lerchianae–Stipetum lessingianae; 34. Eryngiocampestris–Stipetumucrainicae распространены на Донецком кряже и на севере области, однако наибольшее распространение они получили на юго-востоке Ростовской области, в подзоне опустыненных полукустарничково-дерновиннозлаковых степей, где являются ландшафтообразующим типом растительности. Как видно из карты (рис. 6), это наиболее сохранившиеся массивы нераспаханных степей Ергенинской возвышенности и древней долины Западного Маныча. Часто эти степные фитоценозы участвуют в составе комплексов с галофитной растительностью солонцов, выделенной в ассоциации 37. Poobulbosae–Artemisietumpauciflorae.
Сообщества ассоциации 35. Amorioretusae–Cerastietumsyvaschici, а также ассоциации 36. Agropyrinipectinati–Poetumbulbosae, представляющие долинные степи Западного Маныча, хорошо дифференцированы в пространстве.
Таким образом, построение ГИС-моделей растительности существенно дополнило ординацию единиц растительности в пространстве экологических факторов, функциональную модель степного растительного покрова бассейна реки Дон в пределах исследуемой территории Ростовской области.
Заключение
Построенная ГИС-модель отражает экологические закономерности распределения сообществ степной растительности на исследуемой территории и их дифференциацию в пространстве ведущих экологических факторов – общего увлажнения, богатства почв и засоления, литолого-эдафического фактора, в том числе степени кальцефитности почв и почвообразующих пород.
Пелитофитные, гемипсаммофитные, гемипетрофитные и гемигалофитные варианты степного типа растительности оказались четко отграничены от псаммофитной, петрофитной и галофитной растительности в экологическом пространстве.
Реализованные в процессе исследования методы и подходы, представление объема и характера геоботанических данных, применение ординационного градиентного анализа в экологии сообществ и построение ГИС-моделей растительности с целью познания пространственных структурных и функциональных закономерностей, все это в целом позволило создать новую экологическую модель распределения растительного покрова степей бассейна Дона, его климатической обусловленности, географической (территориальной) и экотопической приуроченности.
Таким образом, ординационный подход и построение экологических ГИС-моделей пространственной структуры растительного покрова – это новый подход к оценке фиторазнообразия, основанный на последовательной агрегации количественных критериев, содержащихся в созданной документированной классификации растительности степей бассейна Дона и базе данных геоботанических описаний. Представленные материалы являются инновационным механизмом экологизации сельскохозяйственного производства в регионе [25] и обоснованием крайней необходимости увеличения площади особо охраняемых природных территорий [26], имеют большой потенциал в области охраны природы и экологического мониторинга.
Литература:
- Пузаченко Ю.Г. Математические методы в экологических и географических исследованиях // Учебное пособие. – М.: издательский центр "Академия", 2004. – 416 с.
- Василевич В. И. Что считать естественной классификацией // Философские проблемы современной биологии. М.; Л.: Наука, 1966. С. 177–190.
- Грейг-Смит П. Количественная экология растений. Пер. с англ., М.: Мир, 1967. 359 с.
- Миркин Б.М., Наумова Л.Г. Современное состояние основных концепций науки о растительности. Уфа: Гилем, 2012. 488 с.
- Баянов А. Б., Ямалов С. М., Миркин Б. М. Опыт анализа факторов, определяющих состав луговых сообществ с использованием ординационных подходов (на примере северо-востока республики Башкортостан) // Изв. Самарского науч. центра РАН. 2009. Т. 11, № 1. С. 31–33.
- Мокриевич В. И. Природные кормовые угодья Ростовской области: растительность, хозяйственное использование, мероприятия по улучшению // Флора Нижнего Дона и Северного Кавказа: структура, динамика, охрана, проблемы использования. Ростов-на-Дону, 1991. С. 68.
- Паршутина Л. П. Карта степной растительности Ростовской области // Тез. докл. конф. ''К 100-летию со дня рождения акад. Е. М. Лавренко''. М., 2000. С. 31–33.
- Лавренко Е. М., Карамышева З. В., Никулина Р. И. Степи Евразии. Л.: Наука, 1991. 146 с.
- Карта растительности Ростовской области. М. 1: 800 000 / Под ред. О. Н. Демина, В. И. Мокриевич, Е. М. Цвылев, З. В. Кириленко. Ростов-на-Дону, 2005. 2 л. .
- Полевая геоботаника / Под ред. Е. М. Лавренко А. А. Корчагина. М., Л.: Изд-во АН СССР, 1964. 530 с.
- Зверев А. А. Информационные технологии в исследованиях растительного покрова: Учебное пособие. Томск: ТМЛ-Пресс, 2007. 304 с.
- Демина О.Н. Закономерности распределения и развития растительного покрова степей бассейна Дона (в границах Ростовской области). /О.Н. Демина// Автореферат диссертации на соискание ученой степени доктора биологических наук. Москва, 2011. 50 с.
- Demina O.N. The classification of steppe vegetation of the Don Basin // European Vegetation Survey, 21st Workshop, Vienna (Austria), 24–27 May 2012: 2012. P. 14.
- Hill M. O., Gauch H. G. Detrended correspondence analysis: an improved ordination technique. Vegetatio, 1980, v. 42: P. 47–58.
- Clarke, K. R. Non-parametric multivariate analyses of changes in community structure // Australian Journal of Ecology. 1993 v. 18, P. 117–143.
- McCune B, Mefford M. J. PC-ORD. Multivariate Analysis of Ecological Data, Version 4. MjM Software Design, Glended Beach, Oregon, USA. 1999, 238 p.
- Hill, M. O. TWINSPAN--A FORTRAN program for arranging multivariate data in an ordered two-way table by classification of the individuals and attributes. Ithaca, NY: Ecology and Systematics, Cornell University. 1979.
- Джонгман Р. Г., Тер Брак С. Дж. Ф., Ван Тонгерен О. Ф. Р. Анализ данных в экологии сообществ и ландшафтов. М.: РАСХН, 1999. 306 с.
- Prentice I. C. Non-metric ordination methods in ecology // Journal of Ecology 1977, v. 65: P. 85–94.
- Minchin P. R. An evaluation of the relative robustness of techniques for ecological ordination // Vegetatio, 1987. v. 69: P. 89–107.
- Александрова В. Д. К вопросу о выделении фитоценозов в растительном континууме // Бот. журн. 1965. Т. 50, вып. 9. С. 1248–1267.
- Александрова В. Д. Классификация растительности. Л.: Наука. 1969. 275 с.
- Уиттекер Р. Сообщества и экосистемы / Р. Уиттекер. М.: Прогресс, 1980. 327 с.
- Ермаков Н.Б., Полякова М.А., Попов Д.Ю., Голомовзин В.В. Моделирование пространственной организации растительности горных территорий на основе данных дистанционного зондирования и цифровой модели рельефа// Вычислительные технологии. Том 12. Спец. выпуск. 2007. С. 42-59.
- Савон Д.Ю., Гассий В.В. Сценарий устойчивого развития Ростовской области [Электронный ресурс] // «Инженерный вестник Дона», 2012, №4 ч.1. – Режим доступа: http://www.ivdon.ru/magazine/archive/n4p1y2012/1152 (доступ свободный) – Загл. с экрана. – Яз. рус.
- Поляков, П.В. Критерии рациональности и эффективности использования природных ресурсов [Электронный ресурс] // «Инженерный вестник Дона», 2013, №4. – Режим доступа: http://www.ivdon.ru/magazine/archive/n4y2013/1881 (доступ свободный) – Загл. с экрана. – Яз. рус.